Ruud Toonen
Full Professor
To process information the brain is constantly changing the strength of individual contacts (synapses) between nerve cells. Strict control of synaptic plasticity is important, as dysregulation of this process is often associated with neurological and psychiatric disorders. The main goal of the lab is to advance our understanding of the mechanisms that support synaptic plasticity and their dysfunction in disorders such as Alzheimer’s, epilepsy, schizophrenia and autism to provide novel treatment options and therapeutic targets.
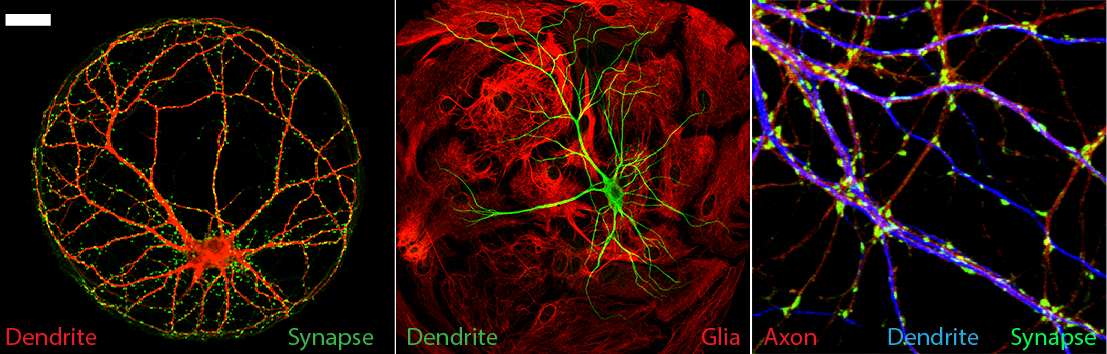
Both pre- and postsynaptic mechanisms contribute to changes in synaptic strength. We focus on presynaptic mechanisms by studying the function of key proteins of the synaptic vesicle fusion machinery, their interactors and downstream effectors in wildtype and disease models. Epilepsy-causing mutations in presynaptic proteins are studied in human IPSC-derived neurons from patients to gain insight into disease-mechanisms for better patient stratification and personalized treatments.
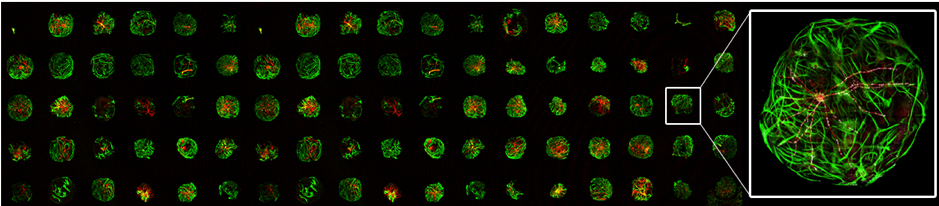 Array of glia micro-islands (green) with human IPSC-derived neurons (red) to study synaptic transmission phenotypes in patient-derived neurons (See Meijer et al., 2019 Cell Reports).
Array of glia micro-islands (green) with human IPSC-derived neurons (red) to study synaptic transmission phenotypes in patient-derived neurons (See Meijer et al., 2019 Cell Reports).
In addition to synaptic vesicles, neurons contain vesicles that store and release many different types of neuromodulatory cargo (neuropeptides and neurotrophins). We study the molecular mechanisms that transport and recruit these vesicles to the plasma membrane, their calcium dependent fusion and the effect of secreted cargo on synaptic plasticity.
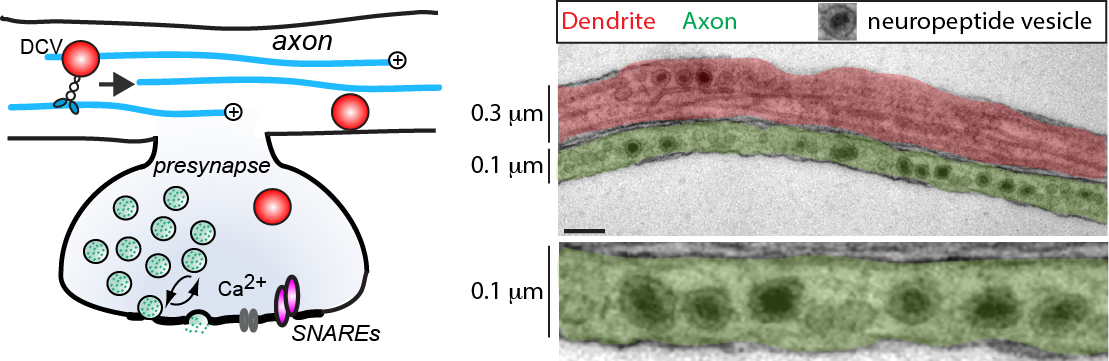
We use wide-field, 2-photon and TIRFM microscopy in combination with electrophysiology to monitor synapse activity and activity-dependent transport, capture and release of secretory vesicles in rodent and human IPSC-derived neurons in vitro and in vivo.
Puntman DC, Arora S, Farina M, Toonen RF#, Verhage M#. Munc18-1 is essential for neuropeptide secretion in neurons (2021) JNeuroSci.
Moro A, van Nifterick A, Toonen RF#, Verhage M#. Dynamin controls neuropeptide secretion by organizing dense-core vesicle fusion sites. (2021) Science Adv. 7(21):eabf0659.
van Westen R, Poppinga J, Díez Arazola R, Toonen RF#, Verhage M#. Neuromodulator release in neurons requires two functionally redundant calcium sensors (2021) PNAS 118(18):e2012137118.
Moro A, Hoogstraaten RI, Persoon CM, Verhage M, Toonen RF. Quantitative analysis of dense-core vesicle fusion in rodent CNS neurons. (2021) STAR Protocols.
Moro A, van Woerden GM, Toonen RF#, Verhage M#. CaMKII controls neuromodulation via neuropeptide gene expression and axonal targeting of neuropeptide vesicles. (2020) Plos Biology. 18(8):e3000826..
Lammertse HCA, van Berkel AA, Iacomino M, Toonen RF, Striano P, Gambardella A, Verhage M, Zara F. Homozygous STXBP1 variant causes encephalopathy and gain-of-function in synaptic transmission. (2019) Brain. 143(2):441-451.
Persoon CM, Hoogstraaten RI, Nassal JP, van Weering JRT, Kaeser PS, Toonen RF#, Verhage M#. The RAB3-RIM Pathway Is Essential for the Release of Neuromodulators. (2019) Neuron. 2019;104(6):1065–1080.
King CR, A A Quadros AR, Chazeau A, Saarloos I, van der Graaf AJ, Verhage M, Toonen RF. Fbxo41 Promotes Disassembly of Neuronal Primary Cilia. (2019) Scientific Reports 9 (1), 8179.
Meijer M, Rehbach K, Brunner JW, Classen JA, Lammertse HCA, van Linge LA, Schut D, Krutenko T, Hebisch M, Cornelisse LN, Sullivan PF, Peitz M, Toonen RF, Brüstle O, Verhage M. A Single-Cell Model for Synaptic Transmission and Plasticity in Human iPSC-Derived Neurons. (2019) Cell Reports 27(7):2199-2211.
Brouwer M, Farzana F, Koopmans F, Chen N, Brunner JW, Oldani S, Li KW, van Weering JRT, Smit AB, Toonen RF#, Verhage M#. SALM1 controls synapse development by promoting F-actin/PIP2-dependent Neurexin clustering. (2019) EMBO J. 2019;38(17):e101289
Persoon CM, Moro A, Nassal JP, Farina M, Broeke JH, Arora S, Dominguez N, van Weering JR, Toonen RF#, Verhage M. Pool size estimations for dense-core vesicles in mammalian CNS neurons.(2018) EMBO J. 2018;37(20).
van Keimpema L, Kooistra R, Toonen RF#, and Verhage M#. CAPS-1 requires its C2, PH, MHD1 and DCV domains for dense core vesicle exocytosis in mammalian CNS neurons. (2017) Scientific Reports 7(1):10817
Arora S, Saarloos I, Kooistra R, van de Bospoort R, Verhage M, Toonen RF.
SNAP-25 gene family members differentially support secretory vesicle fusion. (2017) J. Cell Sci 130(11):1877-1889.
Emperador Melero J, Nadadhur AG, Schut D, Weering JV, Heine VM, Toonen RF#, Verhage M#. Differential Maturation of the Two Regulated Secretory Pathways in Human iPSC-Derived Neurons. (2017) Stem Cell Reports 8(3):659-672.
Schmitz SK, King C, Kortleven C, Huson V, Kroon T, Kevenaar JT, Schut D, Saarloos I, Hoetjes JP, de Wit H, Stiedl O, Spijker S, Li KW, Mansvelder HD, Smit AB, Cornelisse LN, Verhage M, Toonen RF. Presynaptic inhibition upon CB1 or mGlu2/3 receptor activation requires ERK/MAPK phosphorylation of Munc18-1. (2016) EMBO J. 35(11):1236-50
Farina M, van de Bospoort R, He E, Persoon CM, van Weering JR, Broeke JH, Verhage M, Toonen RF. CAPS-1 promotes fusion competence of stationary dense-core vesicles in presynaptic terminals of mammalian neurons. (2015) eLife doi: 10.7554/eLife.05438
Cijsouw T, Weber JP, Broeke JH, Broek JAC, Schut D, Kroon T, Saarloos I, Verhage M and Toonen RF. Munc18-1 redistributes in nerve terminals in an activity- and PKC-dependent manner. (2014) J Cell Biol. 204 (5);759–775
Spangler SA, Schmitz SK, Kevenaar JT, de Graaff E, de Wit H, Demmers J, Toonen RF#, and Hoogenraad CC#. Liprin-α2 promotes the presynaptic recruitment and turnover of RIM1/CASK to facilitate synaptic transmission. (2013) J Cell Biol. 10;201(6):915-28
van de Bospoort R, Farina M, Schmitz SK, de Jong A, de Wit H, Verhage M, Toonen RF.
Munc13 controls the location and efficiency of dense-core vesicle release in neurons. (2012) J Cell Biol. 199(6):883-91.